Научная деятельностьПОЛУЧЕНИЕ И ЭКСПРЕССИЯ КОНСТРУКТА, КОДИРУЮЩЕГО СЛИТНЫЙ БЕЛОК, СОСТОЯЩИЙ ИЗ ОДНОЦЕПОЧЕЧНОГО РЕКОМБИНАНТНОГО АНТИТЕЛА ЛАМЫ, СВЯЗЫВАЮЩЕГО ФНО, И ЦВЕТНОГО БЕЛКА, ФЛУОРЕСЦИРУЮЩЕГО В КРАСНОЙ ЧАСТИ СПЕКТРА.Фактор некроза опухоли (ФНО) является провоспалительным цитокином. В норме, ФНО играет важную роль в защите организма от патогенов, а также участвует в
формировании иммунной системы. Однако, при многих аутоиммунных заболеваниях (в
Устройство генетической конструкции, кодирующей флуоресцентный сенсор TNF – Vhh41-K . Схематическое изображение двухвалентного взаимодействия димера флуоресцентного сенсора с тримером TNF (Ефимов Г.А., Недоспасов С.А., неопубликованные данные)
Прижизненная визуализация областей гиперэкспрессии TNF в модельном аутоиммунном артрите с помощью флуоресцентного сенсора Vhh41-K (Ефимов Г.А., Недоспасов С.А., неопубликованные данные) АНАЛИЗ ЭКСПРЕССИИ TNF В МОДЕЛИ ОСТРОГО ВОСПАЛЕНИЯ С ИСПОЛЬЗОВАНИЕМ РЕПОРТЕРНЫХ МЫШЕЙ С КРАСНЫМ ФЛУОРЕСЦЕНТНЫМ БЕЛКОМРанее в лаборатории ведущего ученого была создана новая линия репортерных мышей (TNF-2A-Кat), содержащих в виде трансгена фрагмент гена TNF с его регуляторными элементами, слитый с геном красного флуоресцентного белка Катюшка (Kat). Таким образом, в геном линии трансгенных TNF-2A-Кat репортерных мышей вставлена генетическая конструкция, позволяющая совместно экспрессировать ген TNF и ген-репортер флуоресцентного белка. Конструкция содержит большинство известных регуляторных элементов гена TNF, что позволяет идентично регулировать экспрессию трансгена и гена TNF клетки. Совместно с НИИ Биомедицинских технологий НижГМА были получены данные по флуоресцентному in vivo имиджингу TNF-2A-Кat репортерных мышей.Так, при сравнении флуоресцентных изображений in vivo было обнаружено, что интенсивность сигнала флуоресценции некоторых тканей TNF-2A-Kat мышей, в частности, кожи, значительно выше по сравнению с интенсивностью сигнала от тканей мышей дикого типа:
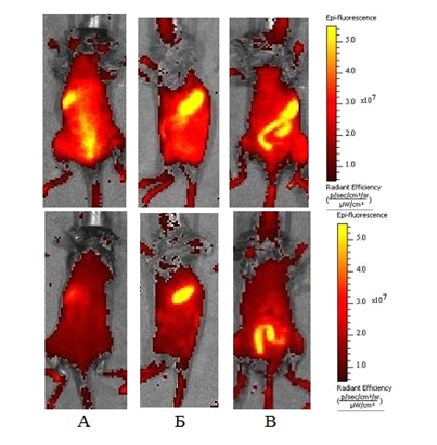
Типичные флуоресцентные изображения, полученные in vivo на мышах TNF-2A- Kat (верхний ряд) и мышах дикого типа (нижний ряд) : А – со стороны спины, Б – с левого бока, В – со стороны живота (яркие желтые пятна являются артефактом аутофлуоресценции). При сравнительном флуоресцентном имиджинге ex vivo внутренних органов TNF-2A-Kat мышей и мышей дикого типа, таких как мышцы, кишечник, желудок, сердце, селезёнка, почки, лимфатические узлы, печень и лёгкие животных. Оценка флуоресценции органов показала, что хотя интенсивность сигнала от кожи, мышц, кишечника, лимфоузлов и легких была выше у TNF-2A-Kat мышей, статистически значимые отличия в интенсивности флуоресценции между двумя группами мышей наблюдались только для кожи и мышечной ткани. Этот результат, полученных на наивных здоровых мышах, разводка которых происходит в SPF виварии, позволил предположить, что активация гена TNF в клетках кожи может быть обусловлена эффектами микробиоты. Эта возможность сейчас исследуется с помощью применения антибиотиков, а также расширением анализа на эмбриональную стадию развития мышей, так как считается, что заселение микробиотой происходит после рождения. Флуоресцентный имиджинг Aldara-индуцированного псориаза у мышей TNF-2A-KatПоскольку репортерные мыши позволили выявить «кожный фенотип» в экспрессии TNF у наивных мышей, возник вопрос об уровне экспрессии этого цитокина в условиях экспериментального воспаления кожи, такого как псориаз. При проведении сравнительного флуоресцентного имиджинга Aldara-индуцированного псориаза у мышей TNF-2A-Kat и мышей дикого типа было отмечено, что после серии последовательных нанесений препарата интенсивность сигнала флуоресценции в зоне заболевания у мышей TNF-2A-Kat значительно увеличивалась по сравнению с контрольным сигналом от здорового участка кожи, тогда как при индукции псориаза у мышей дикого типа подобных различий в интенсивности сигнала флуоресценции между участком кожи с псориазом и участком нормальной кожи не наблюдалось.
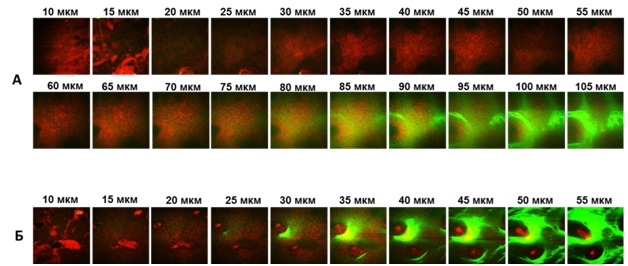
Флуоресцентный имиджинг in vivo и фотографии мыши TNF-2A-Kat (а) и мыши дикого типа (б) в условиях экспериментального псориаза, вызываемого препаратом AldaraТМ. Наблюдение флуоресценции кожи мышей in vivo в динамике после нанесения псориаз-индуцирующего препарата AldaraТМ показало, что несмотря на формирование псориатических бляшек на коже, изменения сигнала, регистрируемого с поверхности у мышей дикого типа, не наблюдалось. Для мышей TNF-2A-Kat было характерно увеличение сигнала флуоресценции в зоне предполагаемой патологии начиная с 3-го дня после нанесения препарата по 7й день анализа, что, вероятно, свидетельствует о повышенном уровне продукции TNF. С помощью установки для многофотонной томографии MPTflex (Jenlab, Германия) были полученыin vivo изображения участков кожи TNF-2A-Kat репортерных мышей после 7 дней нанесения препарата, а также наивных TNF-2A-Kat репортерных мышей, не подвергавшихся воздействиям AldaraTM. Проводили сканирование кожи по глубине с получением Z-стеков с шагом 5 мкм. Анализ изображений показал, что у TNF-2A-Kat репортерных мышей после нанесения препарата AldaraТМ наблюдается значительное утолщение рогового слоя эпидермиса и избыточное количество кератиноцитов по сравнению с интактным участком кожи, что является признаками псориаза:
In vivo изображения псориатического участка кожи TNF-2A-Kat репортерной мыши (А) и здорового участка кожи (Б). Глубина слоя указана над изображениями. Красный канал: автофлуоресценция, двухфотонное возбуждение фемтосекундным лазером 750 нм. Зеленый канал: коллаген, генерация второй гармоники. Регистрация флуоресценции в диапазоне 409-660 нм. СОЗДАНИЕ, ЭКСПРЕССИЯ, ОЧИСТКА И ОЦЕНКА ФУНКЦИОНАЛЬНЫХ СВОЙСТВ БИСПЕЦИФИЧЕСКИХ АНТИТЕЛ НА ОСНОВЕ МИНИ-АНТИТЕЛ А9Одной из основных научных идей проекта, осуществляемого в лаборатории экспериментальной иммунологии, является новый подход к антицитокиновой терапии, основанный на создании более специфических блокаторов TNF, которые будут избирательно нейтрализовать патогенный TNF из конкретных клеточных источников. Для его осуществления на базе уникальных однодоменных антител мозоленогих (ламы, верблюды) были сконструированы биспецифические анти-TNF белки, которые могут нейтрализовать TNF, экспрессируемый макрофагами. Схема генетических конструкций, кодирующих антитело, связывающееся с TNF и макрофагальным маркером F4/80
Экспрессию белков проводили в бактериальной системе (E. coli, штамм Rosetta 2(DE3)pLysS). Очистка осуществлялась с помощью жидкостной аффинной хроматографии на колонках с Ni-NTA Agarose смолой. Затем, для переведения в HEPES-буфер, проводили 3 раунда диализа. Присутствие и концентрацию белка в элюате оценивали по изменению оптической плотности раствора при длине волны 280 нм, с помощью спектрофотометра при 320-340 нм в реакции с трихлоруксусной кислотой, а также методом электрофореза в ПААГ. Для обеспечения стабильности препаратов белков BV1, BV1c и BV1mut были использованы различные химические стабилизаторы, которые согласно нормативам Федерального закона «О лекарственных средствах» являются безопасными в производстве и применении терапевтических препаратов. Максимальная стабильность белков наблюдалась при присутствии в растворе 10% D(+)глюкозы или D(+)сорбитола. Кроме того, добавление к раствору белков 50 мM аргинина и 50 мM глутаминовой кислоты увеличивало их растворимость. Для предотвращения образования спонтанных внутри- и межмолекулярных S-S связей в раствор была введена окислительно-восстановительная пара 25mM L-аскорбиновая кислота/20 mcM CuSO4. Для определения эффективности связывания белков с TNF был использован метод ИФА, который основан на «сэндвич» - гибридизации. Максимальная чувствительность полученных препаратов антител составила около 5 нг/мл рекомбинантного TNF, что свидетельствует о высокой эффективности взаимодействия BV1, BV1c и BV1mut с белком-«мишенью».
Препараты белков BV1, BV1c и BV1mut после очистки и оптимизации стабильности. Оценку способности полученными рекомбинантными антителами блокировать TNF проводили с помощью цитотоксического MTT-теста на клетках линии WEHI-164. По результатам теста все три белка обладали ингибирующей активностью по отношению к TNF, и, в концентрации около 100 нМ, полностью блокировали действие TNF, тем самым сохраняя 100% выживаемость клеток.
Кривые выживаемости клеток линии WEHI-164 после взаимодействия с TNF в концентрации 200ЕД и белков BV1, BV1-c, BV1-mut в разных концентрациях. Данные обрабатывались в программе GraphPadPrism с помощью метода наименьших квадратов. Показаны 95% доверительные интервалы. Исследование BV-1 in vivo на примере модели острой гепатотоксичности, индуцированной введением липополисахарида (ЛПС) и D-галактозамина.Для эксперимента были сформированы три группы мышей, которым за полчаса до инъекции смеси гепатотоксичных агентов ЛПС и D-галактозамина вводили следующие препараты: в качестве положительного контроля 1 группе мышей - препарат мышиного моноклонального антитела F10 в дозе 7 мкг/г веса, группе 2 - исследуемый препарат BV-1 в дозе 5 мкг/г веса, третьей группе (используемой в качестве отрицательного контроля) -фосфатно-солевой буфер (PBS). По результатам теста препарат BV-1 наравне с моноклональным антителом F10 обладает защитным действием, сохраняя 100% выживаемость мышей по сравнению с контрольной группой, которой вводили PBS. В дальнейшем планируется повторить эксперимент для исследуемых белков (BV1, BV1c и BV1mut), чтобы выявить минимальную эффективную дозу.
Кривые и диаграмма выживаемости "гуманизированных" мышей в модели острой гепатотоксичности, индуцированной введением липополисахарида и D-галактозамина. |